题目是“MethylglyoxalForms Diverse Mercaptomethylimidazole Crosslinks with Thiol and Guanidine Pairsin Endogenous Metabolites and Proteins”
通讯作者是芝加哥大学的Raymond E Moellering教授。
他的课题组致力于利用化学蛋白质组学或化学生物学技术研究细胞中与疾病相关的分子机制。
细胞能够通过三羧酸循环、糖酵解通路等途径产生多种活性代谢物,其中一部分可与蛋白质、脂质或核酸上的亲核基团形成稳定的共价修饰,使相应的生物大分子功能发生变化。由细胞中一碳代谢所产生的甲醛在生理浓度下也能够交联蛋白质表面的相邻的氨基酸侧链,形成稳定的亚甲基桥修饰。北京大学的陈鹏课题组最近的工作揭示了一种能够相应甲醛的DNA结合蛋白HxlR的生理机制:甲醛分子通过与HxlR上的Cys11与Lys13交联形成亚甲基桥,使HxlR构象改变并结合DNA;作者利用这一特点构建了能在活细胞与脑组织内检测甲醛的可基因编码荧光探针(Zhu et al, 2021)。另一种被广泛研究的活性代谢物是丙酮醛(methyglyoxal,MGO),它是一种由磷酸丙糖在磷酸消除过程中产生的活性代谢物,它能够与蛋白质的氨基酸侧链(如精氨酸、赖氨酸及半胱氨酸)反应生成多种修饰。Raymond教授课题组此前的工作中报道MGO能够与细胞应激蛋白KEAP1上邻近的精氨酸与半胱氨酸形成稳定的交联修饰(mercaptomethylimidazole crosslinks between cysteine and arginine, MICA)并导致KEAP1二聚,从而激活转录因子Nrf2(Bollong et al, 2018)。除了生物大分子以外,丙酮醛也能够交联细胞内含有胍基与巯基的代谢物,因此系统性研究丙酮醛的反应性质有助于在细胞中鉴定其相关的修饰,在未来这可能助力于对疾病相关的信号转导的研究。
首先,作者在体外将丙酮醛与含有巯基的N-乙酰半胱氨酸(N-acetylcysteine, NAC)与谷胱甘肽(GSH)及含有胍基的精氨酸和甲氨蝶呤(Metformin, Met)分别孵育并通过质谱监测反应动力学。结果表明在反应初始阶段丙酮醛更倾向于与巯基反应,生成硫代半缩醛类修饰(Hemithioacetal);随着反应的进行,硫代半缩醛修饰的含量逐渐减少,取而代之的是与胍基反应后生成的咪唑啉酮修饰(imidazolone)以及交联了胍基与巯基的MICA修饰,这一结果可能是因为生成的硫代半缩醛修饰可逆;作者也发现,较低浓度的丙酮醛能够促进咪唑啉酮类修饰以及MICA修饰的产生。以上结果帮助定义了丙酮醛对巯基与胍基的反应活性,也有助于预测丙酮醛在细胞内可能的代谢产物(见图1)。
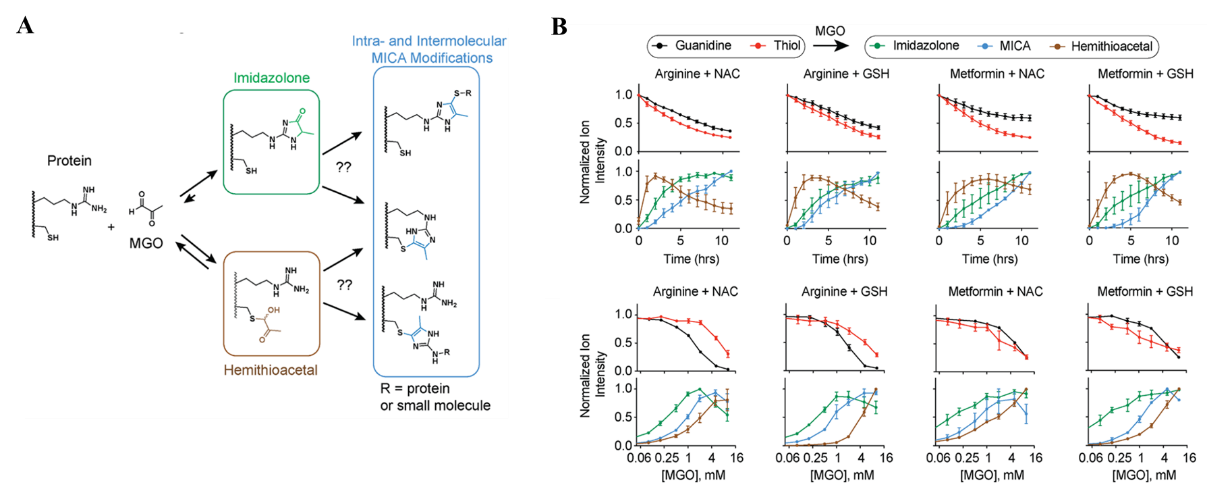
图1. A)丙酮醛与蛋白质侧链胍基与巯基的可能反应产物; B) LC-MS监测丙酮醛与含有巯基或胍基小分子的反应动力学。
接下来为了评估丙酮醛在蛋白质氨基酸侧链上对于巯基与胍基的反应活性,作者在含有巯基与胍基的肽段上探索了丙酮醛反应的活性与热动力学,结果表明丙酮醛倾向于交联同一肽段内的巯基与胍基。作者基于这一结果,提出了丙酮醛形成MICA修饰的两条路径(见图2-C):1)丙酮醛的醛羰基首先被具有亲核性的巯基进攻,再与胍基发生alpha-ketone重排反应生成MICA修饰;2)丙酮醛首先与胍基反应形成咪唑啉酮,巯基再进攻咪唑啉酮的羰基最终脱氢生成MICA修饰。为了验证这两种完全相反的途径,作者先用NAC与精氨酸对肽段进行预处理,再用丙酮醛与其反应。结果表明,用NAC预处理后形成硫代半缩醛的肽段产生MICA修饰的速度更快且产率更高,这一结果证实了起始阶段丙酮醛与巯基之间的反应在MICA修饰的形成过程中起到了主导作用(见图2-A)。作者进一步将丙酮醛与胎牛血清蛋白(Bovine Serum Albumin,BSA)进行孵育,BSA含有一个不形成二硫键的半胱氨酸(Cys58),实验结果表明当BSA与丙酮醛共孵育时,这一位点能够与溶液中的精氨酸形成MICA修饰;而作者并未检测到BSA表面的精氨酸与溶液中的半胱氨酸形成的MICA修饰(见图2-B)。该现象也进一步证实了作者提出的MICA修饰形成路径。
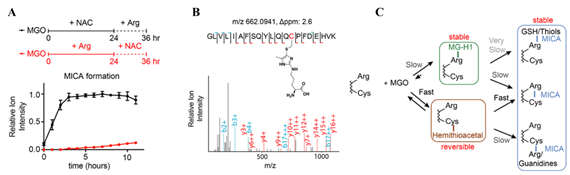
图2. A) NAC能够促进肽段上MICA产物的形成;B) BSA的巯基能够与溶液中的精氨酸形成MICA修饰;C)丙酮醛与巯基和胍基的反应偏好性及机理。
为了进一步探索活细胞内由丙酮醛反应生成的代谢物或修饰,作者建立了一种基于多反应监测的质谱方法,利用代谢组学探究了细胞内丙酮醛与巯基或胍基反应生成的产物。作者发现用一定浓度的丙酮醛处理HeLa细胞后,细胞内能够产生丙酮醛与谷胱甘肽和精氨酸形成的交联产物GSH-Arg-MGO,但作者并未检测到丙酮醛与精氨酸和半胱氨酸反应形成的产物,这可能是因为细胞内游离的半胱氨酸较少。作者还发现在甲氨蝶呤处理的细胞中也能检测到相应的交联产物GSH-Met-MGO (见图3)。
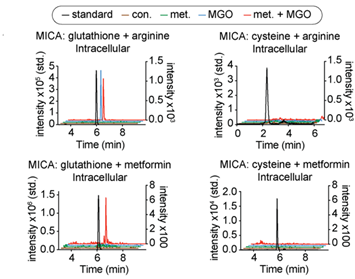
图3.基于代谢组学探究细胞内丙酮醛的交联产物。
丙酮醛分子在细胞内可通过乙二醛酶进行脱毒反应,然而丙酮醛生成的一系列代谢产物及加合物被细胞代谢或排出的机制尚不清楚。作者发现随着丙酮醛浓度的增加,细胞内GSH-Arg-MICA的含量也明显提高,而细胞外并未检测到游离的GSH,这暗示了由丙酮醛产生的交联产物有可能被主动转运到了细胞外(见图4-A)。此前的研究表明一部分谷胱甘肽复合物能够通过由基因ABCC1编码的蛋白MRP1排出细胞外,因此作者假设MRP1可能参与了GSH-Arg-MICA的主动转运过程。为了验证这一假设,作者敲低了HeLa细胞中的MRP1,再用丙酮醛处理后发现GSH-Arg-MICA在细胞内的含量上升,而在细胞外的含量下降(见图4-B),且敲低MRP1并未影响细胞内谷胱甘肽或精氨酸的含量,这一实验结果说明细胞内MICA修饰代谢物,甚至可能是一些含有MICA修饰的蛋白降解产物,能够部分通过MRP1进行主动运输(见图4-C)。
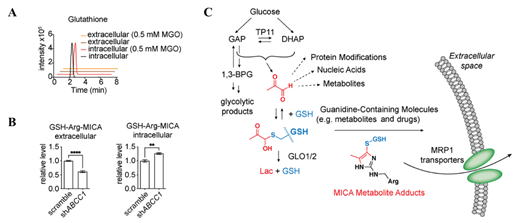
图4. A-B)敲低MRP1探究其在GSH-Arg-MICA主动转运中的的作用;C)丙酮醛在细胞中的来源、代谢以及转运途径。
总之,这篇文章较为系统地研究了丙酮醛在生理环境下对含有巯基及胍基的代谢物或生物大分子的反应活性。文中提出了MICA修饰形成的一种可能的机理,并发现丙酮醛在细胞内的生成的交联产物能够通过蛋白质MRP1转运至胞外,作者认为这可能是细胞内除了乙二醛脱氢酶GLO1/2之外,对丙酮酸进行“脱毒”的又一途径。

